
Photo by CanyonlandsNPS licensed under public domain.
Gardeners are all too familiar with herbivory. Countless times I have been awaiting a bloom to burst only to have the buds nipped off the night before they opened. While this can be devastating for many plant species (not to mention my sanity), for certain plant species, an encounter with a hungry herbivore may actually lead to an increase in reproductive fitness.
Overcompensation theory is the idea that, under certain conditions, plants can respond to herbivore damage by producing more shoots, flowers, and seeds. It goes without saying that when this idea was originally proposed in the late 80's, it was met with its fair share of skepticism. Why would a plant capable of producing more shoots and flowers wait to be damaged to do so? The answer may lie in in the realm of biological trade-offs.
Overcompensation may evolve in lineages that tend to grow in habitats where there is a "predictable" amount of herbivory in any given growing season, perhaps a region where large herbivores migrate through annually. Plants in these habitats may conserve dormant growing tips and valuable resources to be used once herbivory has occurred. Perhaps this also serves as a cue to upregulate antiherbivory compounds in new tissues. The trade-off is that the plants incur a cost in the form of fewer flowers and thus reduced reproduction when herbivory is low or absent.

It could also be that plants are exhibiting two different strategies - one to deal with competition and one to deal with herbivory. If herbivory is low, plants may become more competitive, thus favoring rapid vertical growth of one or a couple shoots. When herbivory is high, rapid vertical growth becomes disadventageous and overcompensatory branching and flowering can provide the higher fitness benefits.
These possibilities are not mutually exclusive. In fact, since the late 80's, experts now believe that overcompensation is not an "either/or" phenomenon but rather a spectrum of possibilities that are dictated by the conditions in which the plants are growing. Certainly overcompensation exists but which conditions favor it and which do not?
Research on scarlet gilia (Ipomopsis aggregata), a biennial native to western North America, suggests that overcompensation comes into play only when environmental conditions are most favorable. Soil nutrients seem to play a role in how well a plant can bounce back following herbivore damage. When resources are high, the results can be quite astounding. Early work on this species showed that under proper conditions, plants that were browsed by upwards of 95% produced 2.4 times as much seed as uneaten control plants. What's more, the resulting seedlings were twice as likely to survive than their uneaten counterparts.
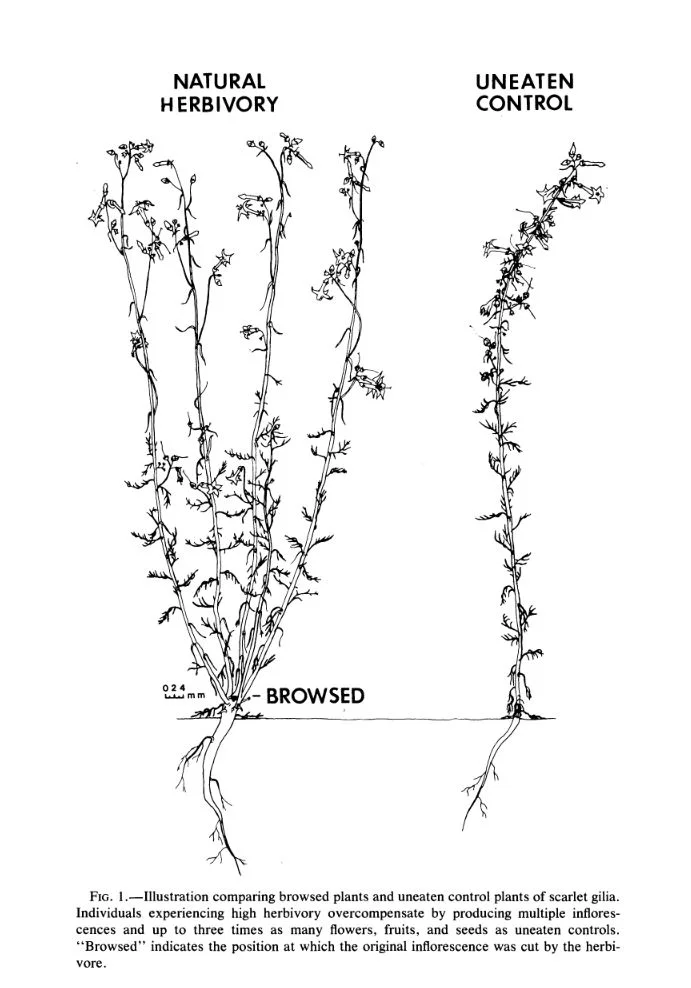
Things change for scarlet gilia growing in poor conditions. Low resource availability appears to place limits on how much any given plant may respond to browsing. Also, herbivory can really hamper flowering time. Because scarlet gilia is pollen limited, anything that can cause a disruption in pollinator visits can have serious consequences for seed set. In at least one study, browsed plants flowered later and received fewer pollinator visits as a result.
More recent work has been able to add more nuance to the overcompensation story. For instance, experiments done on two subspecies of field gentian (Gentianella campestris), add further support to the idea that overcompensation is a matter of trade-offs. They showed that, whereas competition with neighboring plants alone could not explain the benefits of overcompensation, browsing certainly can.

Field gentian (Gentianella campestris). Photo by Joan Simon licensed under CC BY-SA 2.0
Plants growing in environments where herbivory was higher overcompensated by producing more branching, more flowers, and thus more seed, all despite soil nutrients. It appears that herbivory is the strongest predictor of overcompensation for this gentian. What's more, when these data were fed into population models, only the plants that responded to herbivory by overcompensation were predicted to show any sort of population growth in the long term.
Despite all of the interest overcompensation has recieved in the botanical literature, we are only just beginning to understand the biological mechanisms that make it possible. For starters, we know that when a dominant shoot or stem gets damaged or removed, it causes a reduction in the amount of the plant hormone auxin being produced. When auxin is removed, tiny auxiliary buds at the base of the plant are able to break dormancy and begin growing.
Removal of the dominant shoot or stem can also have major impact on the number of chromosomes present in regrowing tissues. Work on Arabidopsis thaliana revealed that when the apical meristem (main growing tip of a vertical stem) was removed, the plant underwent a process called "endoreduplication" in which the cells of the growing tissues actually duplicate their entire genome without undergoing mitosis.

Photo by Hectonichus licensed under CC BY-SA 3.0
Endoreduplication is a complex process with lots of biological significance but in plants it is often associated with stress responses. By duplicating the genomes of these new cells, the plants may be able to adjust more rapidly to their environment. This often manifests in changes to leaf size and shape and an uptick in plant defenses. Thus, plants may be able to fine tune the development of new tissues to overcompensate for browsing. Certainly far more work is needed to understand these mechanisms and their functions in more detail.
Overcompensation is not universal. Nonetheless, it is expected to occur in certain plants, especially those with short life cycles, and under certain environmental conditions, mainly when herbivore pressure and nutrient availability are relatively high. That being said, we still have plenty more to learn about this spectrum of strategies. When does it occur and when does it not? How common is it? What are the biological underpinnings of plants capable of overcompensation? Are some lineages more prone to overcompensation than others? Only more research can say for sure!
Photo Credits: [1] [2] [3] [4] [5]
Further Reading: [1] [2] [3] [4]