
Photo by Fredlyfish4 licensed by CC BY-SA 4.0
Pollination may seem like a strange thing to us humans. Whereas we only require two of us to accomplish reproduction, plants have to utilize a third party. The most familiar cases include insects like bees and butterflies. Unique examples include birds, bats, and even lizards. Many plants forego the need of an animal and instead rely on wind to broadcast copious amounts of pollen into the air in hopes that it will randomly bump into a receptive female organ.
This has worked very well for terrestrial plants but what about their aquatic relatives? Water proves to be quite an obstacle for the methods mentioned above. Some species get around this by thrusting their flowers above the surface but others don't bother. One genus in particular has evolved a truly novel way of achieving sexual reproduction without having to leave its aquatic environment in any way.

Photo by eyeweed licensed by CC BY-NC-ND 2.0
Meet the Vallisnerias. Commonly referred to as tape or eelgrasses, this genus of aquatic plants has been made famous the world over by their use in the aquarium trade. In the wild they grow submerged with their long, grass-like leaves dancing up into the water column. Where they are native, eelgrasses function as an important component of aquatic ecology. Everything from fish and crustaceans all the way up to manatees utilize tape grass beds for both food and shelter. Eelgrasses stabilize stream beds and shorelines and even act as water filters.
All this is quite nice but, to me, the most interesting aspect of Vallisneria ecology is their reproductive strategy. Whereas they will reproduce vegetatively by throwing out runners, it is their method of sexual reproduction that boggles the mind. Vallisneria are dioecious, meaning individual plants produce either male or female flowers. The female flowers are borne on long stalks that reach up to the water surface. Once there they stop growing and start waiting. Because of their positioning, water tension causes a slight depression around the flowers at the surface. The depression resembles a little dimple with a tiny white flower in the center.

A female Vallisneria flower. Photo by eyeweed licensed by CC BY-NC-ND 2.0

Male Vallisneria flowers floating on the water surface. Photo by eyeweed licensed by CC BY-NC-ND 2.0
Male flowers are very different. Much smaller than the female flowers, a single inflorescence can contain thousands of individual male organs. As they mature underwater, the male flowers break off from the inflorescence and float to the surface. Similar to wind pollinated terrestrial plants, Vallisneria use water currents to disperse their pollen. Once at the surface, the tiny male flowers float around like little pollen-filled rafts.
If a male flower floats near the dimple created by a female flower, it will slide down into the funnel-like depression where it will contact with the female flowers. This is how pollination is achieved. Once pollinated, hormonal changes signal the stem of the female flower to begin to coil up like a spring, drawing the developing seeds safely underwater where they will mature. Eventually hundreds of seeds are released into the water currents.
![After pollination, the stem of the female flower coils up, drawing the ripening ovaries safely underwater. Photo by Peter M. Dziuk [source]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1565792376405-64JD8XOEZI9QIVPB430T/vallisneria-americana-15-13.jpg)
After pollination, the stem of the female flower coils up, drawing the ripening ovaries safely underwater. Photo by Peter M. Dziuk [source]
The Vallisneria are incredible aquatic plants. Their bizarre reproductive strategy has ensured that these plants never really have to leave the water. The fact that they can also reproduce vegetatively means that many species are very successful plants. In fact, some species have become noxious invasive weeds where they have been introduced far outside of their native range. If you own these plants in any way, do take the necessary measures to ensure that they never have the chance to become invasive.
Photo Credits: [1] [2] [3] [4]
Further Reading: [1]

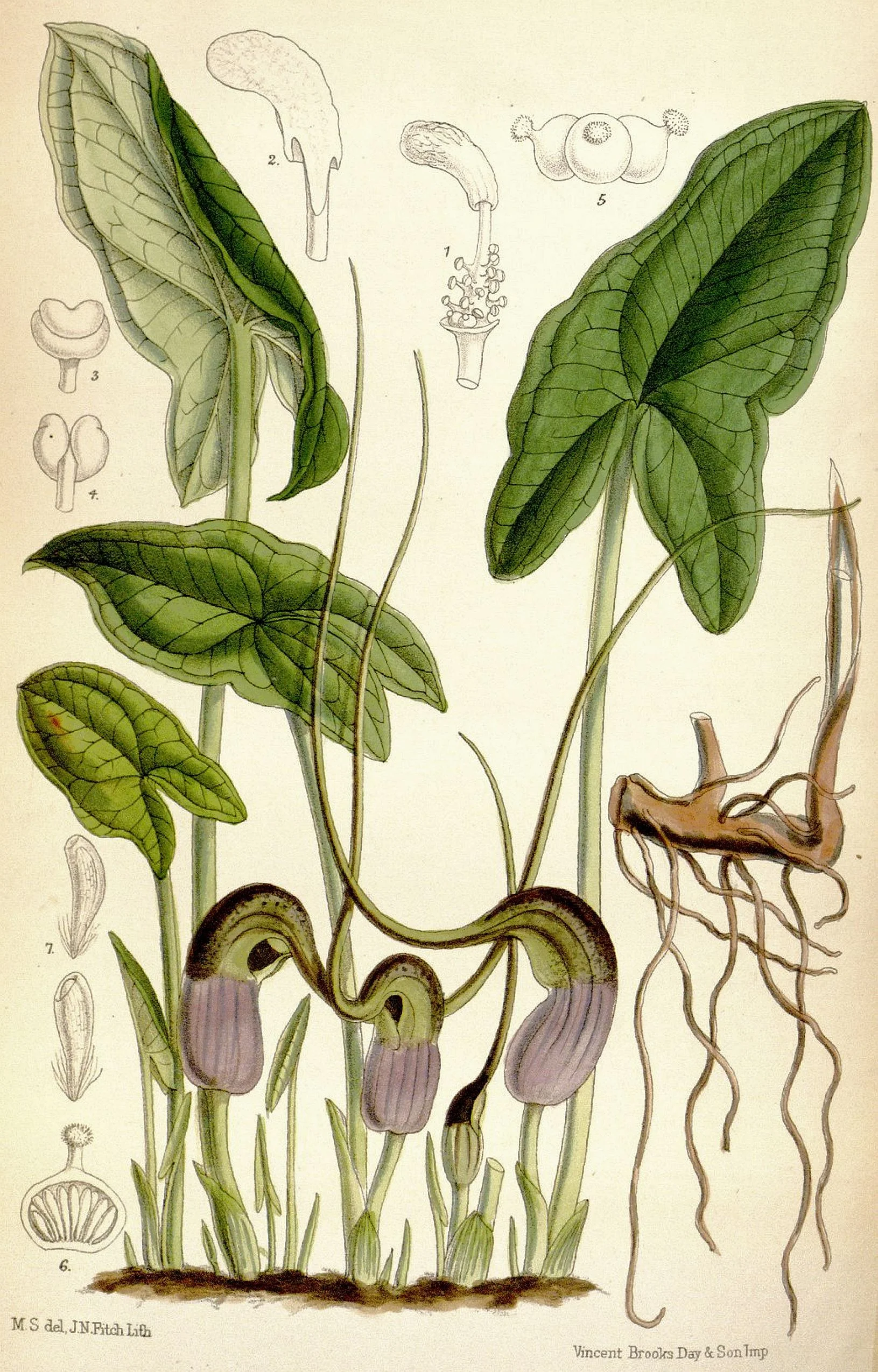
![Anatomy of a mouse plant inflorescence [SOURCE]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1557945730300-PS22LHE82GUKE9OWKEJA/mouse+cross.JPG)



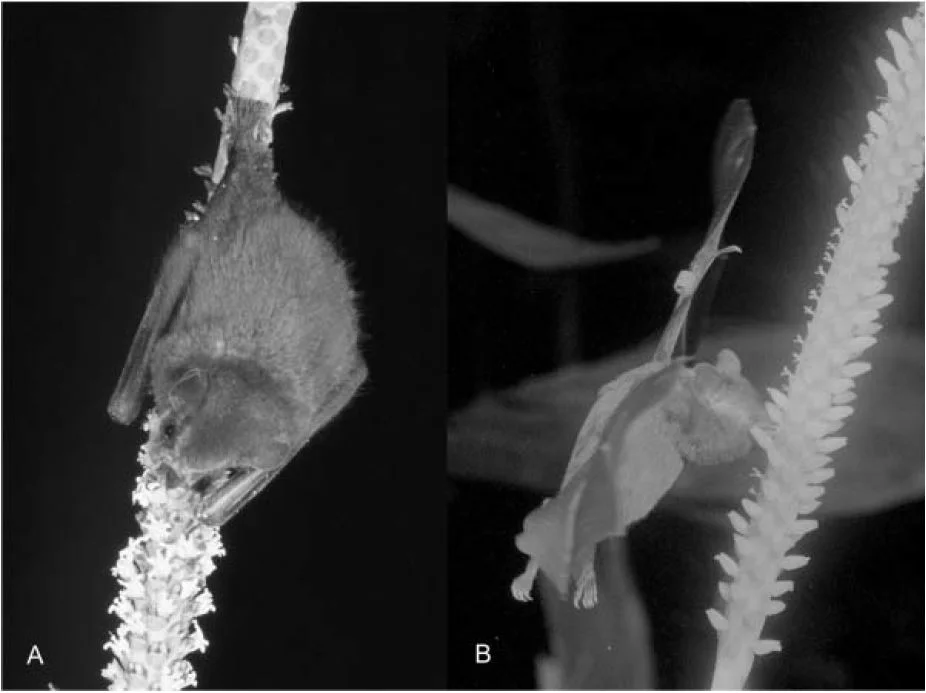
















![Leucospermum arenarium in the field and one of its pollinators, Gerbillurus paeba, feeding on flowers. (A) Pollen presenter contact on G. paeba. (B) G. paeba foraging on L. arenarium [Source]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1537827836454-8MRDJYZTEWG0C9Z67RLJ/pro2.PNG)


![(C) Flowering L. arenarium with dense, mat-forming inflorescences. (D) Geoflorous inflorescences. (E) Pendulous inflorescences above ground level. [Source]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1537827887634-OUACGBZFIHJR2VX4TIL9/pro3.PNG)



















































{kind=link}